FÁRMACOS
BASADOS EN EL ADN

A principios de los años noventa, se comenzó a
difundir el potencial del ADN para inmunizar a las personas.
Los fármacos basados en el ADN están formados por
plásmidos (pequeños anillos de ADN) construidos para transportar un gen hasta
el interior de la célula. Cuando llegan allí, las células sintetizan la
proteína codificada por el gen. Esta proteína provoca la respuesta inmunitaria
que evita una futura infección por parte de ese virus.
Las ventajas de estas vacunas son: su fabricación
mucho más rápida que la de alguna de las vacunas tradicionales puesto que no es
necesario cultivar ni manipular ningún virus vivo, además de no necesitar
refrigeración constante. Otra ventaja es su seguridad, el sistema inmunitario
no percibe los plásmidos como un material extraño ya que no provoca ninguna
respuesta inmunitaria, solo atrae a las células inmunitarias.
Mecanismos
de inducción de respuesta inmune por las vacunas de ADN
Las vacunas de
ADN se basan en la inyección directa en el huésped de un plásmido que codifica
para un antígeno de un patógeno, en lugar del antígeno proteico o del patógeno
atenuado o muerto. Al ser las células del paciente las que producen la
proteína, el antígeno no contiene impurezas, al contrario que en las vacunas
tradicionales, donde en el proceso de purificación pueden quedar trazas de
antibióticos o proteínas del medio de cultivo. La expresión del antígeno dentro
de las células del huésped puede inducir una respuesta inmune completa y
duradera. Esta respuesta incluye anticuerpos, aunque es frecuentemente más
débil que la que se puede obtener con vacunas recombinantes, así como una
activación fuerte y duradera de células T cooperadoras y citotóxicas o de
respuesta celular.
 Un aspecto que
aún no se comprende del todo consiste en la interacción de la vacuna de ADN con
el sistema inmune. Las cantidades del antígeno que se producen cuando se
administra el plásmido están en el orden de los picogramos o nanogramos. Estos
niveles relativamente pequeños de antígeno hacen pensar que la respuesta inmune
tan fuerte y sostenida se debe al tipo de células que capturan el ADN, ya que
es necesario que células especializadas llamadas células presentadoras de
antígeno (CPA) capturen el antígeno lo procesen y lo presenten a otras células
del sistema inmune como como los linfocitos T.
Un aspecto que
aún no se comprende del todo consiste en la interacción de la vacuna de ADN con
el sistema inmune. Las cantidades del antígeno que se producen cuando se
administra el plásmido están en el orden de los picogramos o nanogramos. Estos
niveles relativamente pequeños de antígeno hacen pensar que la respuesta inmune
tan fuerte y sostenida se debe al tipo de células que capturan el ADN, ya que
es necesario que células especializadas llamadas células presentadoras de
antígeno (CPA) capturen el antígeno lo procesen y lo presenten a otras células
del sistema inmune como como los linfocitos T.
Existen tres
mecanismos por el que el antígeno es procesado y presentado al sistema inmune
tras lavacunación con ADN.
a) El ADN desnudo podría ser capturado directamente por las
CPA, éstas células poseen en su superficie una molécula que es conocida como
CMH (Complejo Principal de Histocompatibilidad) de clase II, a través de esta
molécula las CPA presentan el antígeno a los linfocitos T cooperadores,
dependiendo del tipo de célula cooperadora es también el tipo de respuesta que
se activa, para Th1 se activa la respuesta celular, para Th2 se activa la
respuesta humoral y para Th3 se activa la respuesta en mucosas. Se sabe que
éste tipo de células se encuentran presentes en pequeñas cantidades en el
músculo y que constituyen un potente estímulo para el sistema inmune. Tan sólo
se requieren 100 CPAs para activar al sistema inmune, y ésta cantidad no es
difícil de transfectar si consideramos que 8% de las células en la epidermis
corresponden a CPA.
b) El otro mecanismo está relacionado con la adquisición
del ADN por las células musculares y queratinocitos, las cuales no poseen CMH
de clase II, en su lugar poseen CMH de clase I, que ayuda a la activación de
células T de tipo citotóxico con la capacidad de destruir células que se
encuentren infectadas.
c) El tercer mecanismo es una presentación cruzada, es
decir, que a su vez el ADN sea capturado por una célula muscular y por una CPA.

Pero tenía varios inconvenientes. Uno de ellos era
que los plásmidos no llegaban a alcanzar un número suficiente de células y por
lo tanto a producir las proteínas necesarias.
Así que se pensó en nuevos métodos para introducir
a los plásmidos en la célula, en nuevas formas de incrementar la producción de
proteínas. Entre los distintos métodos destacan:
Los parches transdérmicos y otros
sistemas sin aguja que utilizan el aire comprimido para inyectar la vacuna,
introduciendo los plásmidos en la piel, donde abundan los antígenos (un tipo de
célula inmunitaria). En el caso de una inyección con aguja se aplica acto
seguido una electroporación (serie de impulsos eléctricos que provocan la
formación de poros que facilitan la entrada de los plásmidos en las células).
Se modificaron las construcciones gen-plásmido,
retocando la secuencia genética, mejorando la estabilidad y la precisión de los
transcritos génicos en forma de ARN mensajero y acelerando la producción de
proteínas.
Otra mejora consistió en el empleo de adyuvantes
(sustancia que se añade a las vacunas para potenciar la respuesta inmunitaria)
favoreciendo a una mayor producción de células T.
La estrategia basada en el ADN promete la
administración mediante plásmidos de algunos medicamentos y de inmunoterapias
contra el cáncer.
También se suministran genes de factores de
crecimiento para tratar la insuficiencia cardiaca. Otro utiliza un plásmido que
codifica el factor de crecimiento para tratar el retraso de crecimiento.
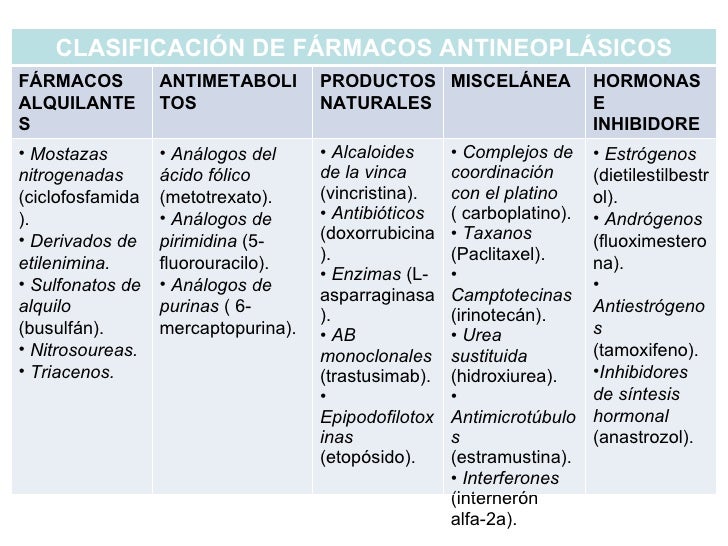
Fármacos que actúan sobre el ADN
Antineoplásicos
Los diferentes fármacos antineoplásicos pueden actuar sobre una o varias
fases del ciclo celular o sobre los mecanismos
de control de la proliferación
celular. La respuesta obtenida se relaciona
directamente con la capacidad
proliferativa de la célula, que está determinada
por el tiempo de duplicación
del tumor. En general, a mayor
proliferación se prevé una mayor respuesta
al tratamiento citostático. En la
evolución del cáncer se van produciendo
nuevas alteraciones genéticas
que provocan una heterogeneidad celular
y, por tanto, unas propiedades
bioquímicas, un tiempo de duplicación
y una respuesta al tratamiento antitumoral
diferentes. Estos mecanismos
están estrechamente ligados a la
aparición de resistencias.
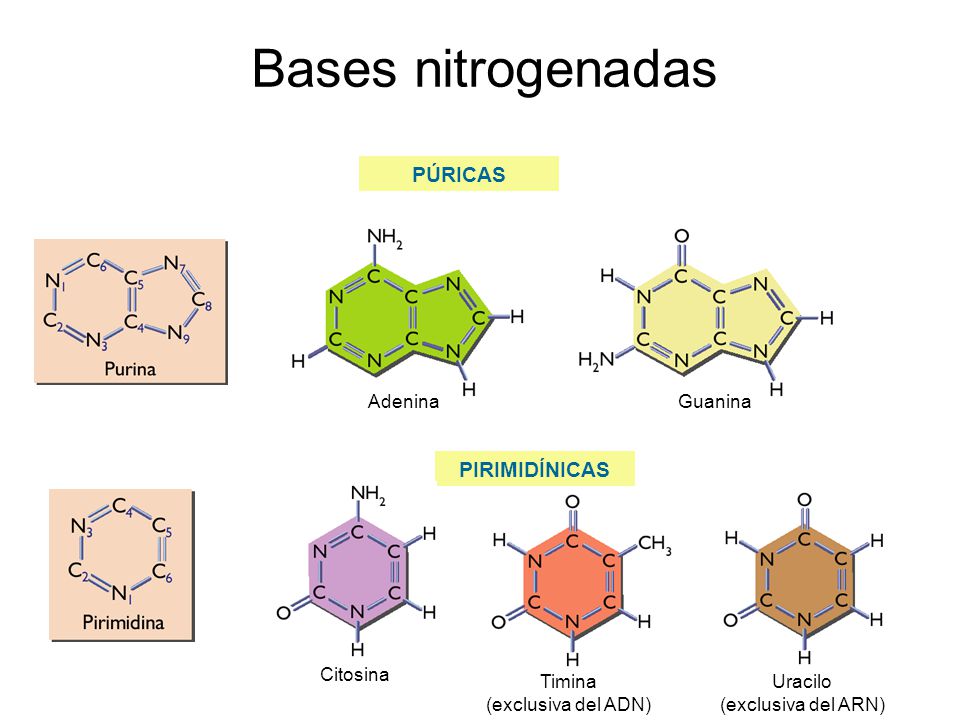
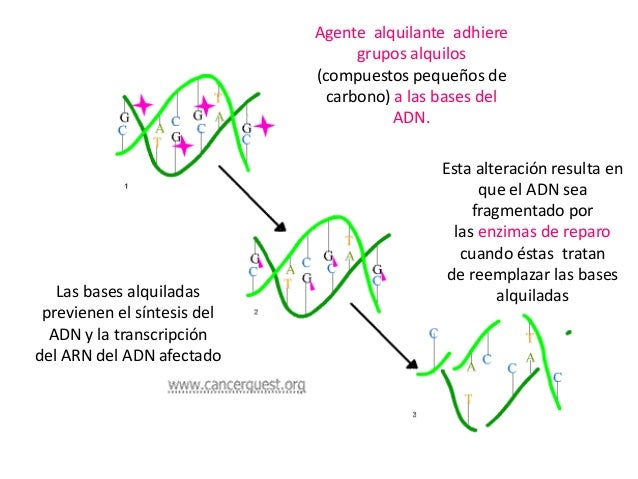
Alquilantes
Mecanismo de acción: Estos fármacos, los más utilizados en
quimioterapia antineoplásica, lesionan
el ADN e interfieren en la replicación
celular. Provocan su acción citotóxica
mediante la formación de enlaces covalentes
entre sus grupos alquilo y diversas
moléculas nucleófilas presentes
en las células, especialmente las bases
nitrogenadas del ADN. De este modo,
bloquean la replicación del ADN celular
y la transcripción del ARN y, por
tanto, la mitosis y la síntesis de proteínas. Ejercen su acción durante todo
el ciclo celular, pero son más activos
sobre las células en rápida división.
Los fármacos de este grupo tienen
en común la toxicidad aguda ocasionada
en la médula ósea en forma de
mielosupresión e inmunosupresión.
Además, afectan a la gametogénesis y
pueden causar esterilidad masculina
permanente; en las mujeres, pueden
reducir el período reproductivo con el
inicio de una menopausia prematura.
También se asocian a un incremento
notable de la incidencia de la leucemia
aguda no linfocítica, sobre todo
cuando se combinan con radioterapia
extensa.
Ejemplos:
Ciclofosfamida. Es el agente alquilante
más utilizado. Se trata de una molécula
inactiva que requiere activación
hepática, con la que da lugar a la fosforamida,
que es el principal metabolito
activo. Por ello se puede administrar
por vía oral y no es vesicante
cuando se administra por vía intravenosa.
 Sus principales efectos tóxicos
son: mielosupresión, alopecia, náuseas
y vómitos. También puede causar
cistitis hemorrágica por la acción de
algunos de sus metabolitos, como la
acroleína, sobre el epitelio de la vejiga;
una ingesta elevada de líquidos
durante 24 o 48 horas puede evitar esta
complicación. Es un fármaco muy
utilizado en oncología y forma parte
de los esquemas de poliquimioterapia,
ya que se ha demostrado su actividad
en diversas neoplasias (leucemias,
linfomas, cáncer de mama, cáncer
de ovario y sarcomas). También
forma parte de los principales regímenes
de inducción previos al trasplante
de médula ósea.
Sus principales efectos tóxicos
son: mielosupresión, alopecia, náuseas
y vómitos. También puede causar
cistitis hemorrágica por la acción de
algunos de sus metabolitos, como la
acroleína, sobre el epitelio de la vejiga;
una ingesta elevada de líquidos
durante 24 o 48 horas puede evitar esta
complicación. Es un fármaco muy
utilizado en oncología y forma parte
de los esquemas de poliquimioterapia,
ya que se ha demostrado su actividad
en diversas neoplasias (leucemias,
linfomas, cáncer de mama, cáncer
de ovario y sarcomas). También
forma parte de los principales regímenes
de inducción previos al trasplante
de médula ósea.
Dosis:
 En adultos:
En adultos:
- Regímenes intermitentes a altas
dosis vía oral o IV (más usual):
La dosis máxima es 40-50 mg / Kg administrada una vez
o en 2-5 días, repetirlo cada 2 4 semanas; estas dosis no son bien toleradas oralmente.
- Vía IV:
10-15 mg/Kg cada 7-10 días o 3-5 mg/Kg 2 veces a la
semana.
- Vía oral diaria continua:
1-5 mg/Kg/día. Las dosis continuas deben
individualizarse según la respuesta del paciente.
En niños: Misma dosis que el adulto.
 Ifosfamida: es un fármaco
análogo a la ciclofosfamida, pero
requiere dosis más altas para conseguir
el mismo efecto antitumoral. Se
administra exclusivamente por vía intravenosa,
siempre con una hidratación
adecuada y mesna como medidas
profilácticas.
Ifosfamida: es un fármaco
análogo a la ciclofosfamida, pero
requiere dosis más altas para conseguir
el mismo efecto antitumoral. Se
administra exclusivamente por vía intravenosa,
siempre con una hidratación
adecuada y mesna como medidas
profilácticas. Dosis:
En adultos:
- Cáncer testicular refractario:
1,2 g/metro cuadrado/día IV en 30 minutos
a 4 horas durante 5 días, o 2 g/metro cuadrado/día durante 3 días consecutivos.
La dosis concurrente de mesna
IV recomendada es el 20% de la dosis de ifosfamida,
administrada 15
minutos antes de la ifosfamida y otra vez
a las 4 y 8 horas. Puede
mezclarse directamente con ifosfamida. Las
últimas 2 dosis de Mesna
 pueden administrarse oralmente en 2 veces
la dosis (por ejemplo,
pueden administrarse oralmente en 2 veces
la dosis (por ejemplo,
cada una el 40% de la dosis de ifosfamida)
si puede asegurarse la
adaptabilidad del paciente y la falta de émesis.
- Alternativamente, vía IV mediante
infusión continua:
5-8 g/metro cuadrado en 24 horas con mesna
añadido en la misma concentración que ifosfamida. Sin embargo, puede ocurrir
nefrotoxicidad más severa con este régimen.
En niños:
- Sarcomas (Sarcoma de Ewing y
osteosarcoma): 1,2 g/metro cuadrado/día IV en 30 minutos
durante 5 días, cada uno con 3
dosis de mesna IV administrada. La dosis
IV de mesna recomendada es el 20% de la dosis de ifosfamida, administrada 15
minutos antes de ifosfamida y otra vez a las 4 y 8 horas. Puede administrarse directamente
con ifosfamida. Las dos últimas dosis de mesna se pueden administrar oralmente
en dos veces la dosis (Ej: cada una el 40% de la dosis de ifosfamida) si puede
asegurarse la adaptabilidad del paciente y la falta de emesis.
 Melfalán: se desarrolló
pensando en el tratamiento del melanoma,
por actuar de forma selectiva
sobre las células tumorales que emplean
de forma activa tirosina. Sin embargo,
en esta neoplasia ha mostrado
escasa actividad. Actualmente, su
principal indicación es el mieloma
múltiple y ha demostrado su eficacia,
a dosis convencionales, en el carcinoma
de ovario y en los linfomas, y a dosis
altas, en el carcinoma de mama y
en la leucemia aguda mieloide.
Melfalán: se desarrolló
pensando en el tratamiento del melanoma,
por actuar de forma selectiva
sobre las células tumorales que emplean
de forma activa tirosina. Sin embargo,
en esta neoplasia ha mostrado
escasa actividad. Actualmente, su
principal indicación es el mieloma
múltiple y ha demostrado su eficacia,
a dosis convencionales, en el carcinoma
de ovario y en los linfomas, y a dosis
altas, en el carcinoma de mama y
en la leucemia aguda mieloide. Dosis:
Dosis:
En adultos:
- 0,25 mg/Kg/día vía oral durante 4 días; o 2-4
mg/día como dosis de
mantenimiento.
16 mg/mL IV cada 2 semanas durante 4 dosis,
luego mensualmente.
Antibióticos
Pocos de los antibióticos que interfieren con las funciones del ADN son útiles en clínica, ya que no pueden discriminar entre ADN de procariotas y eucariotas. Sin embargo, han sido muy valiosos para estudiar diversos aspectos de la biología molecular del ADN.


Actinomicina
– D:
 Mecanismo: El hecho de tener tres anillos
conjugados en un plano le permite intercalarse
entre pares de bases adyacentes de la doble hélice del ADN,
mientras que las dos L-treoninas establecen puentes de H con guaninas del ADN
adyacentes al sitio de intercalación del antibiótico. De esta forma inhibe la replicación del ADN y su
transcripción a ARNm.
Mecanismo: El hecho de tener tres anillos
conjugados en un plano le permite intercalarse
entre pares de bases adyacentes de la doble hélice del ADN,
mientras que las dos L-treoninas establecen puentes de H con guaninas del ADN
adyacentes al sitio de intercalación del antibiótico. De esta forma inhibe la replicación del ADN y su
transcripción a ARNm. Dosis:
Dosis:
En adultos:
- 2 mg/semana IV o 500 mcg/día hasta 5 días,
repetido a las 3 o 4 semanas.
En niños:
- 450 mcg/mL/día, hasta un máximo de 500 mcg/día,
durante 5 días; el ciclo se repite dentro de 3 semanas. La dosis debería
reducirse en presencia de disfunción hepatobiliar.
Mitomicina
 Mecaniso: Al entrar a la célula es convertida
a su forma hidroquinona, que es muy reactiva, funcionando como un agente alquilante bifuncional que
origina entrecruzamientos entre
las dos hebras del ADN. Las consecuencias de ello son: las dos hebras no
pueden separarse durante el intento de replicación, por lo que ésta se detiene.
Mecaniso: Al entrar a la célula es convertida
a su forma hidroquinona, que es muy reactiva, funcionando como un agente alquilante bifuncional que
origina entrecruzamientos entre
las dos hebras del ADN. Las consecuencias de ello son: las dos hebras no
pueden separarse durante el intento de replicación, por lo que ésta se detiene.
A continuación el ADN entrecruzado es atacado
y destruido por las nucleasas de la propia célula.

10-15 mg/metro cuadrado repetida cada 6 semanas si se ha resuelto la toxicidad hematológica.
En regímenes de combinación, se administra en dosis de 5-10 mg/metro cuadrado, repetida cada 4-6 semanas.
Hasta 60 mg/semana se pueden administrar intravesicalmente en cáncer de vejiga.
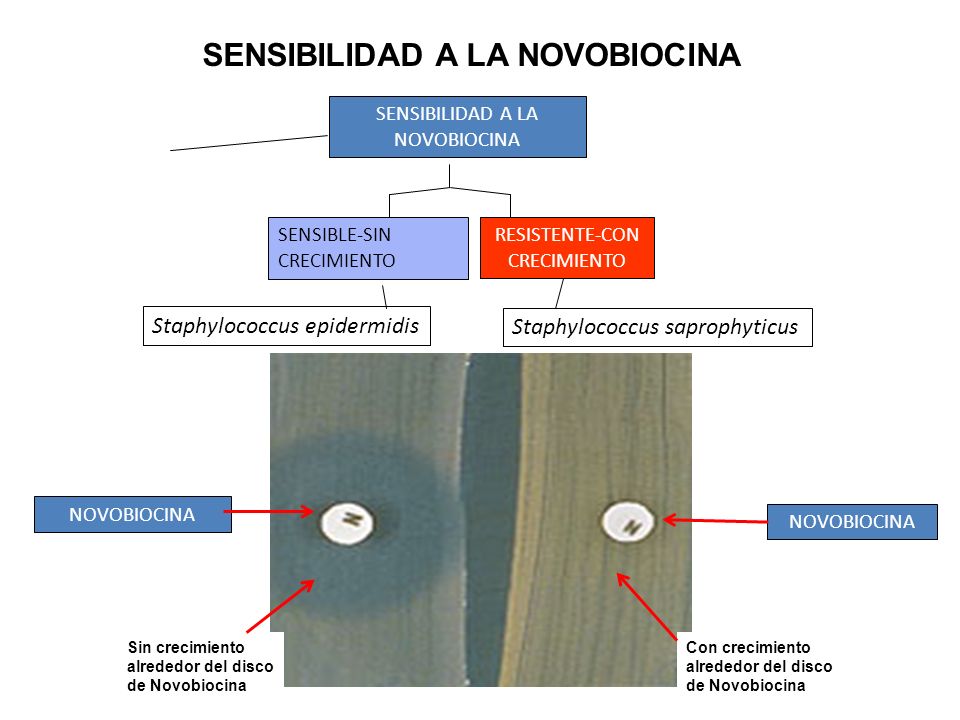
Novobiocina
Mecanismo: Se unen a la subunidad B de las
ADN girasas bacterianas, impidiendo el superenrollamiento negativo del ADN al
competir por el sitio de unión de esta subunidad al ATP.
Dosis:
Infeccions por gérmenes sensibles. Infecciones urinarias resistentes a otros tratamientos. Las dosis recomendadas spn de 250 mg cada 6 horas o 500 mg cada 12 horas. El tratamiento debe continuarse hasta 48 horas después de que la temperatura vuelve a la normalidad.
En los pacientes con insuficiencia renal no se requieren reajustes en las dosis