Lípidos
 Como las
proteínas y los carbohidratos, los lípidos son componentes esenciales de todos
los organismos vivos. Sin embargo, a diferencia de las proteínas y los
carbohidratos, los lípidos tienen estructuras muy variadas. A menudo se definen
como compuestos orgánicos insolubles en agua (o sólo poco solubles), que se
encuentran en los sistemas biológicos. Los lípidos tienen gran solubilidad en
solventes orgánicos no polares. Son hidrofóbicos (no polares) o bien son
anfipáticos (contienen regiones polares y no polares al mismo tiempo).
Como las
proteínas y los carbohidratos, los lípidos son componentes esenciales de todos
los organismos vivos. Sin embargo, a diferencia de las proteínas y los
carbohidratos, los lípidos tienen estructuras muy variadas. A menudo se definen
como compuestos orgánicos insolubles en agua (o sólo poco solubles), que se
encuentran en los sistemas biológicos. Los lípidos tienen gran solubilidad en
solventes orgánicos no polares. Son hidrofóbicos (no polares) o bien son
anfipáticos (contienen regiones polares y no polares al mismo tiempo).
Los
lípidos tienen funciones biológicas diversas:
 ·
Las membranas biológicas contienen una
variedad de lípidos anfipáticos, incluyendo los glicerofosfolípidos y los
esfingolípidos.
·
Las membranas biológicas contienen una
variedad de lípidos anfipáticos, incluyendo los glicerofosfolípidos y los
esfingolípidos.
·
En algunos organismos, la función de los
triacilgliceroles (grasas y aceites) es de moléculas intracelulares de
almacenamiento de energía metabólica.
·
También las grasas suministran aislamiento
térmico y amortiguamiento a los animales.
·
Las ceras en las paredes celulares, en los
exoesqueletos y en la piel protegen a las superficies de algunos organismos.
·
Algunos lípidos tienen funciones muy
especializadas. Por ejemplo, las hormonas esteroides regulan e integran una
variedad de actividades metabólicas en los animales, y los icosanoides participan
en la regulación de la presión sanguínea, en la temperatura corporal y en la
contracción de los músculos lisos en los mamíferos.
·
Los gangliósidos y otros
glicoesfingolípidos están en la superficie celular y pueden participar en el
reconocimiento celular.
v Ácidos grasos
Los ácidos grasos difieren entre sí
en:
-
La longitud de sus colas de hidrocarburo.
-
La cantidad de dobles enlaces
carbono-carbono.
-
Las posiciones de los dobles enlaces en
las cadenas.
-
La cantidad de ramificaciones.

Los ácidos
grasos son una forma de detergente, porque tienen una larga cola hidrofóbica y
una cabeza polar. Como es de esperar, la concentración de ácido graso libre en
las células es muy baja, porque altas concentraciones de ácidos grasos libres
podrían romper las membranas. La mayor parte de los ácidos grasos están
formados por lípidos más complejos. Están unidos a otras moléculas mediante un
enlace de éster en el grupo carboxilo terminal.
Nomenclatura:
ü En la
nomenclatura de IUPAC, al carbono carboxílico se le considera el C-1
y los demás átomos de carbono se numeran en secuencia.
ü En la
nomenclatura común se usan letras griegas para identificar a los átomos de
carbono. El carbono adyacente al carbono carboxílico (C-2, en la nomenclatura
de IUPAC) se le denomina alfa, y los demás carbonos tienen las letras beta, gama,
etcétera. La letra griega 3 (omega) especifica el átomo de carbono más alejado
del grupo carboxilo, cualquiera que sea la longitud de la cola de hidrocarburo.

Los ácidos grasos que no contienen dobles enlaces carbono-carbono se llaman saturados, en tanto que los que tienen al menos un doble enlace carbono-carbono se clasifican como no saturados o insaturados. Los ácidos grasos no saturados que sólo tienen un doble enlace carbono-carbono se llaman monoinsaturados, en tanto que los que tienen dos o más se denominan poliinsaturados. La configuración de los dobles enlaces en los ácidos grasos no saturados es cis, en general. En la nomenclatura de IUPAC, se indican las posiciones de los dobles enlaces mediante el símbolo Δn, en el que el superíndice n, representa el átomo de carbono de menor número en cada par con doble enlace. Los dobles enlaces de la mayor parte de los ácidos grasos poliinsaturados están separados por un grupo metileno, y en consecuencia no están conjugados.
Una
notación taquigráfica para identificar los ácidos grasos usa dos números
separados por dos puntos (:); el primero indica la cantidad de átomos de
carbono en el ácido graso, y el segundo, la cantidad de dobles enlaces
carbono-carbono, con sus posiciones indicadas como superíndices después de una
letra griega Δ. Ejemplos:
-
Palmitato
se escribe como 16:0
-
Oleato
como 18:1 Δ 9
-
Araquidonato
como 20:4 Δ 5,8,11,14.

También se pueden especificar los ácidos grasos no
saturados por la localización del último doble enlace de la cadena. Algunos
ácidos grasos se llaman:
ü
ɯ-3
(por ejemplo 18:3 Δ
9,12,15)
ü
ɯ-6
(por ejemplo, 18:2 Δ9,12)
ü
ɯ-9
(por ejemplo 18:1 Δ9).

Las
propiedades físicas de los ácidos grasos saturados y no saturados son muy
variadas. Típicamente, los ácidos grasos saturados son sólidos céreos a
temperatura ambiente (22 °C), en tanto que los ácidos grasos no saturados son
líquidos a esta temperatura.
La longitud de la cadena de hidrocarburo
en un ácido graso, y su grado de instauración, influyen sobre el punto de
fusión.
-
A medida que aumentan las longitudes de
las colas de hidrocarburo, también aumentan los puntos de fusión de los ácidos
grasos saturados. A medida que se alarga la cadena, aumenta la cantidad de
interacciones de Van der Waals entre las colas vecinas de hidrocarburo, por lo
que se requiere más energía para separar las interacciones.
La
abundancia relativa de determinados ácidos grasos varía de acuerdo con el tipo
de organismo, su órgano (en los organismos multicelulares) y su fuente
alimenticia.
Los ácidos
grasos más abundantes en los animales suelen ser: oleato (18:1), palmitato
(16:0) y estearato (18:0).
Los
mamíferos requieren ciertos ácidos grasos poliinsaturados en su dieta, que no
pueden sintetizar, como linoleato (18:2), abundante en los aceites vegetales, y
linolenato (18:3), que abunda en aceites vegetales y de pescado. Esos ácidos
grasos se llaman ácidos grasos esenciales. Los mamíferos pueden sintetizar
otros ácidos grasos poliinsaturados si tienen un suministro adecuado de
linoleato y linolenato.
Triacilgliceroles
Los ácidos grasos son combustibles
metabólicos importantes, en especial en los mamíferos.
 Como los átomos de carbono de los
ácidos grasos están más reducidos que los de las proteínas o los carbohidratos,
la oxidación de los ácidos grasos produce más energía (~37 kJ g–1) que la
oxidación de proteínas o carbohidratos (~16 kJ g–1 cada uno). En general, los
ácidos grasos se almacenan en forma de lípidos neutros llamados triacilgliceroles
o triglicéridos.
Como los átomos de carbono de los
ácidos grasos están más reducidos que los de las proteínas o los carbohidratos,
la oxidación de los ácidos grasos produce más energía (~37 kJ g–1) que la
oxidación de proteínas o carbohidratos (~16 kJ g–1 cada uno). En general, los
ácidos grasos se almacenan en forma de lípidos neutros llamados triacilgliceroles
o triglicéridos.
Los triacilgliceroles están
formados por tres residuos de acilo graso esterificados con glicerina, un
azúcar alcohol de tres carbonos. Los triacilgliceroles son muy hidrofóbicos.
En consecuencia, a diferencia de
otros carbohidratos, se pueden almacenar en células en forma anhidra, esto es,
las moléculas no están solvatadas por agua, lo cual ocuparía espacio y añadiría
masa, reduciendo la eficiencia del almacenamiento de energía.
Glicerofosfolípidos
Los lípidos más abundantes en la
mayor parte de las membranas son los glicerofosfolípidos (que también se
llaman fosfoglicéridos) tienen un soporte de glicerol. Los glicerofosfolípidos
más sencillos, los fosfatidatos, consisten en dos grupos acilo graso
esterificados en el C-1 y C-2 del 3-fosfato de glicerol. Hay tres grupos acilo
graso esterificados con la glicerina en los triacilgliceroles, en tanto que
sólo hay dos grupos acilo graso (R1 y R2) en los glicerofosfolípidos. La
propiedad distintiva de los grupos acilo (R1 y R2) de los glicerofosfolípidos
es la presencia de un grupo fosfato en el C-3 del soporte del glicerol. Las
estructuras de los glicerofosfolípidos se pueden dibujar como derivados del
L-glicerol 3-fosfato (o 3-fosfato de L-glicerol), con el sustituyente en el C-2
a la izquierda en una proyección de Fischer. Para simplificar, con frecuencia
se mostrarán esos compuestos como estructuras no referidas a su estereoquímica.

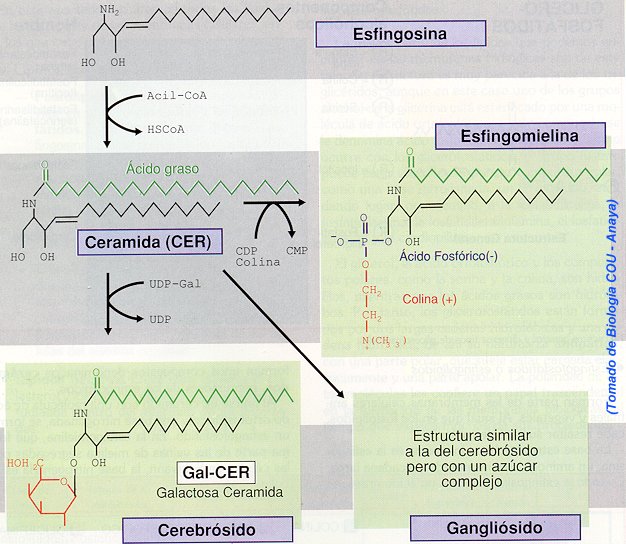
Esfingolípidos
Después de los glicerofosfolípidos,
los lípidos más abundantes en las membranas vegetales y animales son los esfingolípidos.
En los mamíferos tienen abundancia especial en tejidos del sistema nervioso
central. La mayor parte de las bacterias no tienen esfingolípidos. El respaldo
estructural de los esfingolípidos es la esfingosina (trans-4-esfingenina),
un alcohol no ramificado de C18, con un doble enlace trans entre el C-4
y C-5, un grupo amino en el C-2 y grupos hidroxilo en el C-1 y C-3.
La ceramida está formada por un grupo acilo
graso unido al grupo amino del C-2 en la esfingosina, por un enlace de amida. Las
ceramidas son los precursores metabólicos de todos los esfingolípidos. Las tres
grandes familias de esfingolípidos son las esfingomielinas, los cerebrósidos y
los gangliósidos.
En las esfingomielinas, la
fosfocolina está unida al grupo hidróxido en el C-1 de una ceramida, existen en
las membranas plasmáticas de la mayor parte de las células de mamíferos, y son
componente principal de las vainas de mielina que rodean a ciertas células
nerviosas.
Los cerebrósidos son
glicoesfingolípidos que contienen un residuo de monosacárido unido a un enlace b-glicosídico
al C-1 de una ceramida. Los galactocerebrósidos, llamados también
galactosilceramidas, tienen un solo residuo de b-D-galactosilo como grupo de
cabeza polar. Los galactocerebrósidos abundan en el tejido nervioso, y forman
casi el 15% de los lípidos en las vainas de mielina. Muchos otros tejidos en los
mamíferos contienen glucocerebrósidos, ceramidas con un grupo b-D-glucosilo en
la cabeza. En algunos glicoesfingolípidos, una cadena lineal hasta de tres
residuos más de monosacárido está unida a la mitad de galactosilo o glucosilo
de un cerebrósido.
Los gangliósidos son
glicoesfingolípidos más complejos, donde las cadenas de oligosacárido que
contienen ácido N-acetilneuramínico (NeuNAc) están unidas a una ceramida.
El NeuNAc es un derivado acetilado del carbohidrato ácido siálico, y forma los grupos
de cabeza en gangliósidos aniónicos.
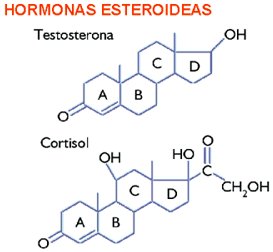
Esteroides
Los
esteroides son una tercera clase de lípidos que se encuentran en las membranas
de los eucariotas, y muy rara vez en las bacterias. Los esteroides, junto con
las vitaminas lipídicas y los terpenos, se clasifican como isoprenoides porque
sus estructuras se relacionan con la molécula de isopreno, de cinco carbonos.
Los esteroides contienen cuatro anillos fundidos, tres de seis carbonos
identificados como A, B y C, y un anillo D de cinco carbonos. La estructura
anular característica se deriva del escualeno. Los sustituyentes del sistema
anular casi plano pueden apuntar hacia abajo (la configuración a) o hacia
arriba (la configuración b).
El
esteroide colesterol es el componente importante de las membranas plasmáticas animales,
pero sólo se encuentra rara vez en las plantas, y nunca en los procariotas,
protistas u hongos. En realidad, el colesterol es un esterol porque
tiene un grupo hidroxilo en el C-3.
El
colesterol se acumula con frecuencia en depósitos lipídicos (placas) en las
paredes de los vasos sanguíneos. Se han identificado esas placas en las
enfermedades cardiovasculares, que pueden provocar ataques al corazón. Muchas personas
limitan su ingestión de colesterol. A pesar de su implicación en enfermedades
cardiovasculares, el colesterol tiene participación esencial en la bioquímica
de los mamíferos. Es sintetizado por las células de mamíferos. No sólo es un
componente de ciertas membranas, sino también un precursor esencial de las
hormonas esteroides y de las sales biliares.

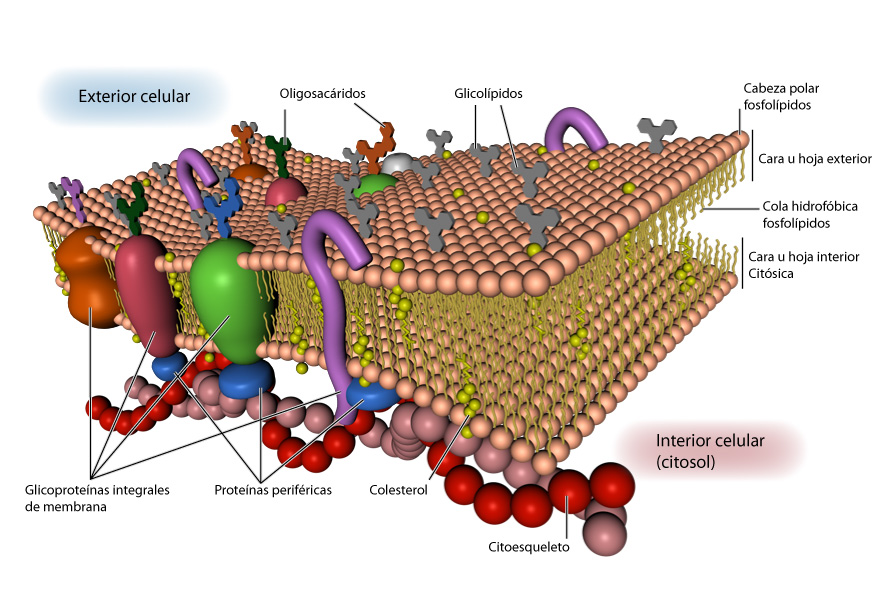
Membranas biológicas
Las membranas biológicas no sólo
son barreras pasivas contra la difusión. Tienen una gran variedad de funciones
complejas. Algunas proteínas contenidas en las membranas sirven como bombas
selectivas que controlan en forma estricta el transporte de iones y de
moléculas pequeñas que entran y salen de la célula. Las membranas también son
responsables de generar y mantener la concentración de gradientes de protones, esenciales
para la producción de ATP. Los receptores en las membranas reconocen señales extracelulares
y las comunican al interior de la célula.
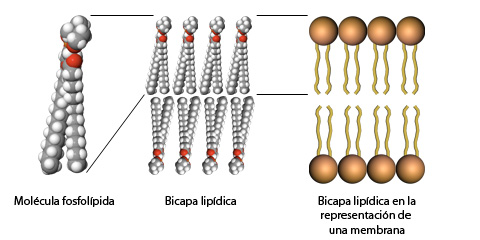
Bicapa
lipídica:
-
Las
bicapas lipídicas son el principal componente estructural de todas las
membranas biológicas, incluyendo membranas plasmáticas y membranas internas de
células eucariotas. Las interacciones no covalentes.
-
Una
bicapa lipídica tiene en forma típica de 5 a 6 nm de espesor, y consiste en dos
hojas o monocapas (que también se llaman hojillas). En cada hoja, los grupos
polares de cabeza en los lípidos anfipáticos están en contacto con el medio
acuoso, y las colas no polares de hidrocarburos apuntan hacia el interior de la
bicapa. La formación espontánea de bicapas lipídicas es impulsada por las
interacciones hidrofóbicas.
No hay comentarios:
Publicar un comentario